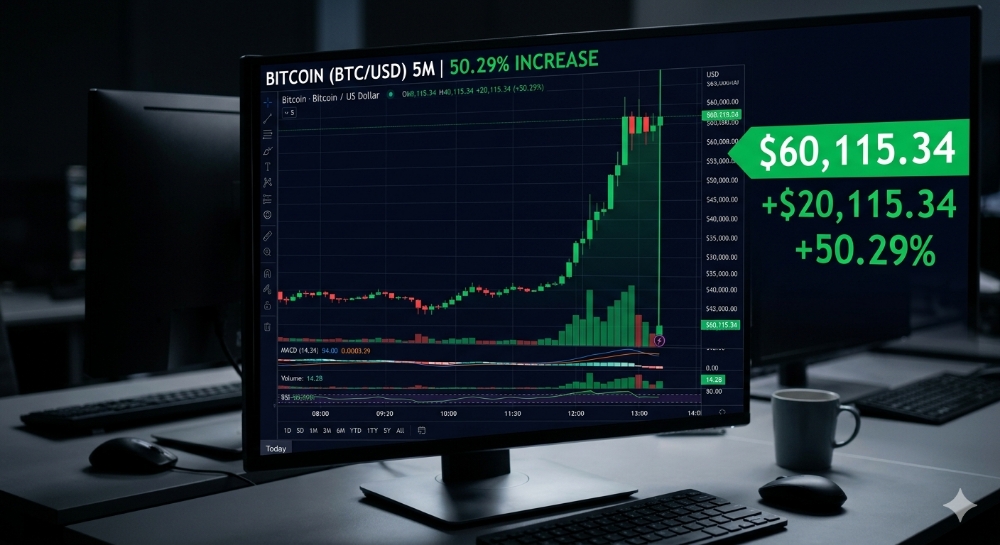
Animals, plants, and microbes have evolved diverse forms of sexual reproduction, which is the mixing of genetic material from different parents. FIGURE 1 Sexual reproduction is remark- ably diverse. (A) The leopard slug (Limax maximus) is a hermaphrodite. Mating pairs hang from a mucus thread while copulat- ing (B) Clownfishes (genus Amphiprion) are sequential hermaphrodites that first mature as male and later change sex to female. (C) The cactus Epithelantha micromeris has flowers that do not open. It is one of the few species that reproduce almost entirely by self-fertiliza- fion. (D) Bacteria, such as these E. coli, use con- jugation and othermechanisms to exchange genetic material.
FIGURE 1 Sexual reproduction is remark- ably diverse. (A) The leopard slug (Limax maximus) is a hermaphrodite. Mating pairs hang from a mucus thread while copulat- ing (B) Clownfishes (genus Amphiprion) are sequential hermaphrodites that first mature as male and later change sex to female. (C) The cactus Epithelantha micromeris has flowers that do not open. It is one of the few species that reproduce almost entirely by self-fertiliza- fion. (D) Bacteria, such as these E. coli, use con- jugation and othermechanisms to exchange genetic material.
Males and females are distinguished by the size of the gametes they make: males make small gametes, and females big ones. Many species are hermaphroditic. In species with separate males and females, sexual dimorphism ranges from minimal to extreme.
▪ Sexual selection, which is selection caused by competition among individuals of the same sex for mates, leads to the evolution of exaggerated secondary sexual traits that increase mating success but usually decrease survival.
▪ Sexual selection acts on males much more often than on females. ▪ Males can often increase their fitness by mating with more females, but females typically do not benefit from mating with more males.
- Some unusual species show sex role reversal. Here sexual selection acts on females because the operational sex ratio is female-biased: more females are available to mate than males. FIGURE 2 Sex role reversal. (A) Two female red phalaropes (Phalaropus fulicarius) fight over the smaller, duller-plumaged male on their breeding ground. In contrast to most birds, female phalaropes court males, which care for the eggs and young in their nests. (B) A male Austra- lian seahorse (Hippocompus breviceps) giving birth from his pouch. Males choose which courting females will lay eggsin. ther pouches.
FIGURE 2 Sex role reversal. (A) Two female red phalaropes (Phalaropus fulicarius) fight over the smaller, duller-plumaged male on their breeding ground. In contrast to most birds, female phalaropes court males, which care for the eggs and young in their nests. (B) A male Austra- lian seahorse (Hippocompus breviceps) giving birth from his pouch. Males choose which courting females will lay eggsin. ther pouches.
One of the two major modes of sexual selection is male-male competition, which occurs by
- male combat,
- sperm competition,
- infanticide,
- other mechanisms.
Male – Male Combat FIGURE 3 Males of many animals have evolved horns and other weapons that they use to fight with each other for reproductive access to females. (A) Male red deer (Cervus elaphus) fighting during the breeding season. Successful males guard groups of females with which they mate. (B) Male stag beetles (Lucanus cervus) fighting. Males defend the sites where females lay their eggs, and mate with females when they arrive to lay.
FIGURE 3 Males of many animals have evolved horns and other weapons that they use to fight with each other for reproductive access to females. (A) Male red deer (Cervus elaphus) fighting during the breeding season. Successful males guard groups of females with which they mate. (B) Male stag beetles (Lucanus cervus) fighting. Males defend the sites where females lay their eggs, and mate with females when they arrive to lay.
Different Strategies FIGURE 4 These isopods (Paracerceis sculpta) live on and inside sponges. The large alpha male guards a harem of females by blocking the entrance to the cavity in which they live. A medium-sized beta male (left) and a small gamma male (right) try to gain access to the females by tricking the alpha male. The beta male imitates a female, while the small gamma male seeks to reach the females by slipping past the alpha male.
FIGURE 4 These isopods (Paracerceis sculpta) live on and inside sponges. The large alpha male guards a harem of females by blocking the entrance to the cavity in which they live. A medium-sized beta male (left) and a small gamma male (right) try to gain access to the females by tricking the alpha male. The beta male imitates a female, while the small gamma male seeks to reach the females by slipping past the alpha male.
Sperm Competition FIGURE 5 A male damselfly removes the sperm of a female’s previous mates before depositing his own. (A) In this photo of mating damselflies (Calopteryx splendens), the blue individual is the male and the gray individual the female. When they mate, claspers on the end of the male’s abdomen grasp the female behind her head. She then curls her abdomen so that the opening of her oviduct (at the end of her abdomen) makes contact with his penis (on his thorax, near the base of his wings). They then copulate. (B) A scoop-like structure on the male’s penis removes the sperm of males that copulated with the female earlier. A clump of sperm adheres to the structure, at bottom. (B courtesy of J. Waage.)
FIGURE 5 A male damselfly removes the sperm of a female’s previous mates before depositing his own. (A) In this photo of mating damselflies (Calopteryx splendens), the blue individual is the male and the gray individual the female. When they mate, claspers on the end of the male’s abdomen grasp the female behind her head. She then curls her abdomen so that the opening of her oviduct (at the end of her abdomen) makes contact with his penis (on his thorax, near the base of his wings). They then copulate. (B) A scoop-like structure on the male’s penis removes the sperm of males that copulated with the female earlier. A clump of sperm adheres to the structure, at bottom. (B courtesy of J. Waage.)
Infanticide FIGURE 5 This male lion has killed a cub after displacing the cub’s father and other adult males in the group that he recently joined. Male infanticide has also been described in many other species of mammals.
FIGURE 5 This male lion has killed a cub after displacing the cub’s father and other adult males in the group that he recently joined. Male infanticide has also been described in many other species of mammals.
The second major mode of sexual selection is female choice.
- Female mating preferences evolve as the result of direct benefits that females receive from their mates, pleiotropic effects of preference genes, and the good genes mechanism.
- Some preferences result from perceptual biases and apparently did not originate by either direct benefits or good genes.
- A final mechanism for the evolution of female preferences is Fisher's runaway process. FIGURE 6 Males of some species provide direct benefits to females. This female Mormon cricket (Anabrus simplex) will eat the large white spermatophore her mate has placed in her genital opening. (Courtesy of John Alcock.)
FIGURE 6 Males of some species provide direct benefits to females. This female Mormon cricket (Anabrus simplex) will eat the large white spermatophore her mate has placed in her genital opening. (Courtesy of John Alcock.) FIGURE 7 Males of the Andean cock-ofthe-rock (Rupicola peruvianus) perform mating displays together on leks. Females visit the lek, mate with the male of their choice, and then leave to raise the offspring alone. There is no opportunity for direct benefits to females in lek-breeding species.
FIGURE 7 Males of the Andean cock-ofthe-rock (Rupicola peruvianus) perform mating displays together on leks. Females visit the lek, mate with the male of their choice, and then leave to raise the offspring alone. There is no opportunity for direct benefits to females in lek-breeding species.
▪ Sexual selection on plants favors flowers that increase pollinator visitation, production of greater quantities of pollen, and pollen that outcompetes other pollen in fertilizing ovules.
- The sex ratio can evolve in many species. In most situations, selection favors producing equal numbers of males and females. FIGURE 8 Selection usually favors a sex ratio with nearly equal numbers of males and females. In this example, females initially produce more daughters than sons, so the sex ratio is biased toward an excess of females. Generation 1: a mutation occurs in one female (red) that causes her to produce only sons. Generation 2: the mutation is now carried by the son of the female that carried the mutation in generation 1. There are only three males, and each of them will be the father to one-third of the individuals in the next generation. Generation 3: the mutation has increased in frequency because each male in the previous generation is father to one-third of the population, and each of his offspring has a probability of 1/2 of inheriting the mutation. Later generations: the mutation will continue to spread until there are equal numbers of males and females. At that equilibrium, the mutation will be more common in males because females with the mutation produce only sons, while males with the mutation produce both sons and daughters that carry the mutation.
FIGURE 8 Selection usually favors a sex ratio with nearly equal numbers of males and females. In this example, females initially produce more daughters than sons, so the sex ratio is biased toward an excess of females. Generation 1: a mutation occurs in one female (red) that causes her to produce only sons. Generation 2: the mutation is now carried by the son of the female that carried the mutation in generation 1. There are only three males, and each of them will be the father to one-third of the individuals in the next generation. Generation 3: the mutation has increased in frequency because each male in the previous generation is father to one-third of the population, and each of his offspring has a probability of 1/2 of inheriting the mutation. Later generations: the mutation will continue to spread until there are equal numbers of males and females. At that equilibrium, the mutation will be more common in males because females with the mutation produce only sons, while males with the mutation produce both sons and daughters that carry the mutation. FIGURE 9 The fig wasp Tetrapus costaricensis (inset) is highly sexually dimorphic. Females lay their eggs inside a fig. When the offspring hatch, the males fight each other, and the winner then mates with the females inside the fig—many or all of whom are his sisters.
FIGURE 9 The fig wasp Tetrapus costaricensis (inset) is highly sexually dimorphic. Females lay their eggs inside a fig. When the offspring hatch, the males fight each other, and the winner then mates with the females inside the fig—many or all of whom are his sisters. FIGURE 10 A mathematical model predicts that natural selection will favor female fig wasps to produce few sons if only one female lays her eggs in a fig, but to produce increasingly equal numbers of sons and daughters as the number of females that lay eggs in the fig increases (shown by the blue curve). In nature, the number of females that enter a fig to lay eggs varies. When only a single female lays her eggs in a fig, fewer than 25 percent of her offspring are male. With increasing numbers of females, the sex ratio tends toward equal numbers of sons and daughters (red dots), in good agreement with the theoretical prediction.
FIGURE 10 A mathematical model predicts that natural selection will favor female fig wasps to produce few sons if only one female lays her eggs in a fig, but to produce increasingly equal numbers of sons and daughters as the number of females that lay eggs in the fig increases (shown by the blue curve). In nature, the number of females that enter a fig to lay eggs varies. When only a single female lays her eggs in a fig, fewer than 25 percent of her offspring are male. With increasing numbers of females, the sex ratio tends toward equal numbers of sons and daughters (red dots), in good agreement with the theoretical prediction.  FIGURE 11Asexual reproduction is very rare among animals and plants. Examples include (A) bdelloid rotifers (Philodina roseola is shown here), (B) some whiptail lizards (Aspidoscelis uniparens is shown here), and (C) the common dandelion (Taraxacum officinale).
FIGURE 11Asexual reproduction is very rare among animals and plants. Examples include (A) bdelloid rotifers (Philodina roseola is shown here), (B) some whiptail lizards (Aspidoscelis uniparens is shown here), and (C) the common dandelion (Taraxacum officinale).
➢ Recombination gives sexual reproduction several advantages that compensate for its disadvantages and thereby explain why it is so common.
➢ The Red Queen hypothesis suggests that sex is favored in changing environments.
➢ Recombination is also favored because it reduces selective interference, which is a general term that includes clonal interference, the ruby-in-the rubbish effect, and Muller's ratchet.
✓ Inbreeding depression frequently causes the evolution of mechanisms that prevent self- fertilization and mating between close relatives.